what type of antigen presenting cells are unable to phagocytize other cells?
The cells of the immune system originate in the os marrow, where many of them besides mature. They then migrate to guard the peripheral tissues, circulating in the claret and in a specialized organisation of vessels called the lymphatic system.
i-1. The white blood cells of the allowed arrangement derive from precursors in the bone marrow
All the cellular elements of blood, including the ruby blood cells that transport oxygen, the platelets that trigger claret clotting in damaged tissues, and the white blood cells of the immune system, derive ultimately from the same progenitor or precursor cells—the hematopoietic stem cells in the os marrow. As these stalk cells can give rise to all of the different types of blood cells, they are often known every bit pluripotent hematopoietic stem cells. Initially, they requite rise to stalk cells of more express potential, which are the immediate progenitors of red claret cells, platelets, and the two main categories of white blood cells. The different types of blood jail cell and their lineage relationships are summarized in Fig. 1.3. Nosotros shall exist concerned hither with all the cells derived from the common lymphoid progenitor and the myeloid progenitor, autonomously from the megakaryocytes and red claret cells.

Figure 1.3
All the cellular elements of blood, including the lymphocytes of the adaptive allowed system, arise from hematopoietic stem cells in the bone marrow. These pluripotent cells divide to produce two more specialized types of stalk cells, a common lymphoid (more...)
The myeloid progenitor is the precursor of the granulocytes, macrophages, dendritic cells, and mast cells of the immune system. Macrophages are 1 of the three types of phagocyte in the immune system and are distributed widely in the body tissues, where they play a disquisitional part in innate amnesty. They are the mature grade of monocytes, which circulate in the blood and differentiate continuously into macrophages upon migration into the tissues. Dendritic cells are specialized to have up antigen and display information technology for recognition by lymphocytes. Young dendritic cells migrate from the blood to reside in the tissues and are both phagocytic and macropinocytic, ingesting large amounts of the surrounding extracellular fluid. Upon encountering a pathogen, they quickly mature and drift to lymph nodes.
Mast cells, whose blood-borne precursors are not well defined, also differentiate in the tissues. They mainly reside near pocket-size blood vessels and, when activated, release substances that affect vascular permeability. Although best known for their role in orchestrating allergic responses, they are believed to play a role in protecting mucosal surfaces against pathogens.
The granulocytes are and then chosen because they take densely staining granules in their cytoplasm; they are as well sometimes called polymorphonuclear leukocytes considering of their oddly shaped nuclei. In that location are iii types of granulocyte, all of which are relatively curt lived and are produced in increased numbers during immune responses, when they leave the blood to migrate to sites of infection or inflammation. Neutrophils, which are the 3rd phagocytic prison cell of the immune organization, are the about numerous and most important cellular component of the innate allowed response: hereditary deficiencies in neutrophil part lead to overwhelming bacterial infection, which is fatal if untreated. Eosinophils are thought to be of import chiefly in defense against parasitic infections, because their numbers increase during a parasitic infection. The office of basophils is probably similar and complementary to that of eosinophils and mast cells; nosotros shall talk over the functions of these cells in Chapter 9 and their function in allergic inflammation in Chapter 12. The cells of the myeloid lineage are shown in Fig. 1.4.
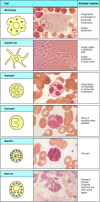
Figure 1.4
Myeloid cells in innate and adaptive immunity. Cells of the myeloid lineage perform various of import functions in the immune response. The cells are shown schematically in the left column in the form in which they will exist represented throughout the rest (more...)
The mutual lymphoid progenitor gives ascension to the lymphocytes, with which most of this book will exist concerned. In that location are two major types of lymphocyte: B lymphocytes or B cells, which when activated differentiate into plasma cells that secrete antibodies; and T lymphocytes or T cells, of which there are 2 master classes. One course differentiates on activation into cytotoxic T cells, which kill cells infected with viruses, whereas the second grade of T cells differentiates into cells that actuate other cells such as B cells and macrophages.
Most lymphocytes are small, featureless cells with few cytoplasmic organelles and much of the nuclear chromatin inactive, every bit shown by its condensed land (Fig. 1.5). This appearance is typical of inactive cells and it is not surprising that, as recently as the early 1960s, textbooks could describe these cells, now the central focus of immunology, every bit having no known function. Indeed, these small lymphocytes have no functional activity until they encounter antigen, which is necessary to trigger their proliferation and the differentiation of their specialized functional characteristics.

Figure 1.5
Lymphocytes are mostly small-scale and inactive cells. The left console shows a low-cal micrograph of a pocket-size lymphocyte surrounded by red claret cells. Note the condensed chromatin of the nucleus, indicating little trans-criptional activity, the relative absenteeism (more...)
Lymphocytes are remarkable in being able to mount a specific immune response against virtually any foreign antigen. This is possible because each individual lymphocyte matures bearing a unique variant of a prototype antigen receptor, so that the population of T and B lymphocytes collectively comport a huge repertoire of receptors that are highly diverse in their antigen-binding sites. The B-jail cell antigen receptor (BCR) is a membrane-jump form of the antibody that the B cell volition secrete subsequently activation and differentiation to plasma cells. Antibody molecules every bit a class are known as immunoglobulins, usually shortened to Ig, and the antigen receptor of B lymphocytes is therefore too known every bit membrane immunoglobulin (mIg). The T-cell antigen receptor (TCR) is related to immunoglobulin merely is quite distinct from it, every bit information technology is especially adapted to detect antigens derived from foreign proteins or pathogens that have entered into host cells. Nosotros shall draw the structures of these lymphocyte antigen receptors in detail in Capacity 3, 4, and 5, and the way in which their diversity of binding sites is created equally lymphocytes develop in Chapter 7.
A third lineage of lymphoid cells, called natural killer cells, lack antigenspecific receptors and are part of the innate immune arrangement. These cells circulate in the blood as large lymphocytes with distinctive cytotoxic granules (Fig. 1.6). They are able to recognize and impale some abnormal cells, for example some tumor cells and virus-infected cells, and are thought to exist of import in the innate immune defense against intracellular pathogens.

Figure one.six
Natural killer (NK) cells. These are large granular lymphocyte-like cells with important functions in innate immunity. Although lacking antigen-specific receptors, they tin can detect and attack sure virus-infected cells. Photograph courtesy of N. Rooney (more...)
one-2. Lymphocytes mature in the bone marrow or the thymus
The lymphoid organs are organized tissues containing large numbers of lymphocytes in a framework of nonlymphoid cells. In these organs, the interactions lymphocytes make with nonlymphoid cells are important either to lymphocyte development, to the initiation of adaptive immune responses, or to the sustenance of lymphocytes. Lymphoid organs tin can be divided broadly into cardinal or primary lymphoid organs, where lymphocytes are generated, and peripheral or secondary lymphoid organs, where adaptive immune responses are initiated and where lymphocytes are maintained. The central lymphoid organs are the bone marrow and the thymus, a large organ in the upper breast; the location of the thymus, and of the other lymphoid organs, is shown schematically in Fig. 1.7.

Figure i.7
The distribution of lymphoid tissues in the torso. Lymphocytes arise from stalk cells in bone marrow, and differentiate in the key lymphoid organs (yellowish), B cells in bone marrow and T cells in the thymus. They migrate from these tissues and are carried (more than...)
Both B and T lymphocytes originate in the os marrow only only B lymphocytes mature there; T lymphocytes migrate to the thymus to undergo their maturation. Thus B lymphocytes are so-chosen because they are bone marrow derived, and T lymphocytes because they are thymus derived. Once they accept completed their maturation, both types of lymphocyte enter the bloodstream, from which they drift to the peripheral lymphoid organs.
1-3. The peripheral lymphoid organs are specialized to trap antigen, to let the initiation of adaptive immune responses, and to provide signals that sustain recirculating lymphocytes
Pathogens can enter the body by many routes and set upwardly an infection anywhere, only antigen and lymphocytes will eventually encounter each other in the peripheral lymphoid organs—the lymph nodes, the spleen, and the mucosal lymphoid tissues (see Fig. 1.7). Lymphocytes are continually recirculating through these tissues, to which antigen is also carried from sites of infection, primarily inside macrophages and dendritic cells. Inside the lymphoid organs, specialized cells such as mature dendritic cells display the antigen to lymphocytes.
The lymph nodes are highly organized lymphoid structures located at the points of convergence of vessels of the lymphatic arrangement, an all-encompassing system of vessels that collects extracellular fluid from the tissues and returns it to the claret. This extracellular fluid is produced continuously by filtration from the blood, and is called lymph. The vessels are lymphatic vessels or lymphatics (see Fig. 1.7). Afferent lymphatic vessels drain fluid from the tissues and also conduct antigen-begetting cells and antigens from infected tissues to the lymph nodes, where they are trapped. In the lymph nodes, B lymphocytes are localized in follicles, with T cells more diffusely distributed in surrounding paracortical areas, also referred to as T-jail cell zones. Some of the B-cell follicles include germinal centers, where B cells are undergoing intense proliferation after encountering their specific antigen and their cooperating T cells (Fig. one.8). B and T lymphocytes are segregated in a similar fashion in the other peripheral lymphoid tissues, and we shall run into that this organisation promotes the crucial interactions that occur betwixt B and T cells upon encountering antigen.
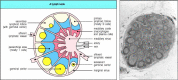
Figure one.viii
Organization of a lymph node. As shown in the diagram on the left, a lymph node consists of an outermost cortex and an inner medulla. The cortex is composed of an outer cortex of B cells organized into lymphoid follicles, and deep, or paracortical, areas (more...)
The spleen is a fist-sized organ simply behind the stomach (see Fig. 1.7) that collects antigen from the blood. It as well collects and disposes of senescent red claret cells. Its organization is shown schematically in Fig. 1.nine. The bulk of the spleen is composed of cherry lurid, which is the site of cherry blood cell disposal. The lymphocytes surround the arterioles entering the organ, forming areas of white lurid, the inner region of which is divided into a periarteriolar lymphoid sheath (PALS), containing mainly T cells, and a flanking B-cell corona.
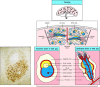
Figure 1.9
Organisation of the lymphoid tissues of the spleen. The schematic at height right shows that the spleen consists of red pulp (pink areas in the top panel), which is a site of red blood jail cell destruction, interspersed with lymphoid white pulp. An enlargement of (more...)
The gut-associated lymphoid tissues (GALT), which include the tonsils, adenoids, and appendix, and specialized structures called Peyer's patches in the pocket-sized intestine, collect antigen from the epithelial surfaces of the gastrointestinal tract. In Peyer's patches, which are the well-nigh important and highly organized of these tissues, the antigen is collected by specialized epithelial cells called multi-fenestrated or M cells. The lymphocytes grade a follicle consisting of a large cardinal dome of B lymphocytes surrounded past smaller numbers of T lymphocytes (Fig. 1.ten). Like but more diffuse aggregates of lymphocytes protect the respiratory epithelium, where they are known as bronchial-associated lymphoid tissue (BALT), and other mucosa, where they are known simply as mucosal-associated lymphoid tissue (MALT). Collectively, the mucosal immune system is estimated to contain every bit many lymphocytes as all the rest of the body, and they class a specialized set of cells obeying somewhat different rules.
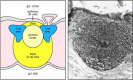
Figure 1.ten
Organisation of typical gut-associated lymphoid tissue. As the diagram on the left shows, the bulk of the tissue is B cells, organized in a large and highly active domed follicle. T cells occupy the areas between follicles. The antigen enters across a specialized (more...)
Although very different in appearance, the lymph nodes, spleen, and mucosal-associated lymphoid tissues all share the same basic architecture. Each of these tissues operates on the same principle, trapping antigen from sites of infection and presenting it to migratory small lymphocytes, thus inducing adaptive immune responses. The peripheral lymphoid tissues likewise provide sustaining signals to the lymphocytes that do not encounter their specific antigen, so that they continue to survive and recirculate until they encounter their specific antigen. This is important in maintaining the correct numbers of circulating T and B lymphocytes, and ensures that just those lymphocytes with the potential to respond to strange antigen are sustained.
one-iv. Lymphocytes broadcast between blood and lymph
Small B and T lymphocytes that have matured in the bone marrow and thymus but have not yet encountered antigen are referred to every bit naive lymphocytes. These cells circulate continually from the blood into the peripheral lymphoid tissues, which they enter by squeezing between the cells of capillary walls. They are and then returned to the blood via the lymphatic vessels (Fig. 1.11) or, in the case of the spleen, return directly to the blood. In the event of an infection, lymphocytes that recognize the infectious amanuensis are arrested in the lymphoid tissue, where they proliferate and differentiate into effector cells capable of combating the infection.
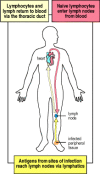
Effigy 1.xi
Circulating lymphocytes encounter antigen in peripheral lymphoid organs. Naive lymphocytes recirculate constantly through peripheral lymphoid tissue, hither illustrated as a lymph node backside the knee, a popliteal lymph node. Here, they may run into their (more...)
When an infection occurs in the periphery, for case, large amounts of antigen are taken upwardly by dendritic cells which then travel from the site of infection through the afferent lymphatic vessels into the draining lymph nodes (see Fig. 1.11). In the lymph nodes, these cells display the antigen to recirculating T lymphocytes, which they besides assist to activate. B cells that meet antigen as they migrate through the lymph node are also arrested and activated, with the assistance of some of the activated T cells. In one case the antigen-specific lymphocytes have undergone a period of proliferation and differentiation, they exit the lymph nodes as effector cells through the efferent lymphatic vessel (see Fig. 1.8).
Because they are involved in initiating adaptive immune responses, the peripheral lymphoid tissues are non static structures just vary quite dramatically depending upon whether or not infection is nowadays. The diffuse mucosal lymphoid tissues may appear in response to infection so disappear, whereas the architecture of the organized tissues changes in a more defined manner during an infection. For example, the B-cell follicles of the lymph nodes expand as B lymphocytes proliferate to form germinal centers (see Fig. 1.viii), and the entire lymph node enlarges, a phenomenon familiarly known equally swollen glands.
Summary
Immune responses are mediated past leukocytes, which derive from precursors in the bone marrow. A pluripotent hematopoietic stalk jail cell gives rising to the lymphocytes responsible for adaptive immunity, and also to myeloid lineages that participate in both innate and adaptive immunity. Neutrophils, eosinophils, and basophils are collectively known equally granulocytes; they circulate in the blood unless recruited to act as effector cells at sites of infection and inflammation. Macrophages and mast cells complete their differentiation in the tissues where they act as effector cells in the front end line of host defence force and initiate inflammation. Macrophages phagocytose leaner, and recruit other phagocytic cells, the neutrophils, from the blood. Mast cells are exocytic and are thought to orchestrate the defense confronting parasites as well as triggering allergic inflammation; they recruit eosinophils and basophils, which are besides exocytic. Dendritic cells enter the tissues as immature phagocytes where they specialize in ingesting antigens. These antigen-presenting cells later on migrate into lymphoid tissue. In that location are two major types of lymphocyte: B lymphocytes, which mature in the os marrow; and T lymphocytes, which mature in the thymus. The os marrow and thymus are thus known as the central or chief lymphoid organs. Mature lymphocytes recirculate continually from the bloodstream through the peripheral or secondary lymphoid organs, returning to the bloodstream through the lymphatic vessels. Most adaptive immune responses are triggered when a recirculating T cell recognizes its specific antigen on the surface of a dendritic jail cell. The iii major types of peripheral lymphoid tissue are the spleen, which collects antigens from the blood; the lymph nodes, which collect antigen from sites of infection in the tissues; and the mucosal-associated lymphoid tissues (MALT), which collect antigens from the epithelial surfaces of the body. Adaptive allowed responses are initiated in these peripheral lymphoid tissues: T cells that encounter antigen proliferate and differentiate into antigen-specific effector cells, while B cells proliferate and differentiate into antibody-secreting cells.
Source: https://www.ncbi.nlm.nih.gov/books/NBK27092/
0 Response to "what type of antigen presenting cells are unable to phagocytize other cells?"
Post a Comment